
· For research use only. Not for human consumption.
Hypothalamic pituitary axis growth peptide research sits at the heart of preclinical endocrinology because this axis controls how the body produces growth hormone (GH) — and the research peptides that interact with it each hit a different, well-defined point in that control system (PubMed search: hypothalamic-pituitary axis GH secretagogues). Think of the axis as a thermostat loop: one part of the brain (the hypothalamus) sends a "produce GH" signal down to the pituitary gland, the pituitary releases GH into the bloodstream, and then a downstream hormone called IGF-1 feeds back to tell the brain to ease off. Research peptides let scientists turn individual steps in this loop up or down, one at a time, so they can study what each step actually does.
Why does any of this matter for experimental design? Because choosing the wrong compound for a given question is like using a screwdriver when you need a wrench. A GHRH analog (like tesamorelin) copies the brain's "produce GH" signal directly at the pituitary. A growth hormone secretagogue (like ipamorelin) works through a completely different receptor — one that responds to a gut hormone called ghrelin — and activates GH release through a separate internal pathway. These two classes can even be combined, because they work through different mechanisms and their effects add up to more than either alone.
This overview maps the key parts of the axis, explains how published research uses growth peptides to probe each part, and covers what compound purity is needed for reliable assay data.
TL;DR: Hypothalamic pituitary axis growth peptide research breaks down into three main classes: compounds that mimic the brain's direct "produce GH" signal (GHRH analogs), compounds that activate a separate ghrelin-related receptor (GHS-R1a agonists), and compounds that reduce the brain's "slow down" signal (somatostatin pathway modulators). Researchers use these as precision tools to study each step of the feedback loop in cell and animal models. For research use only.
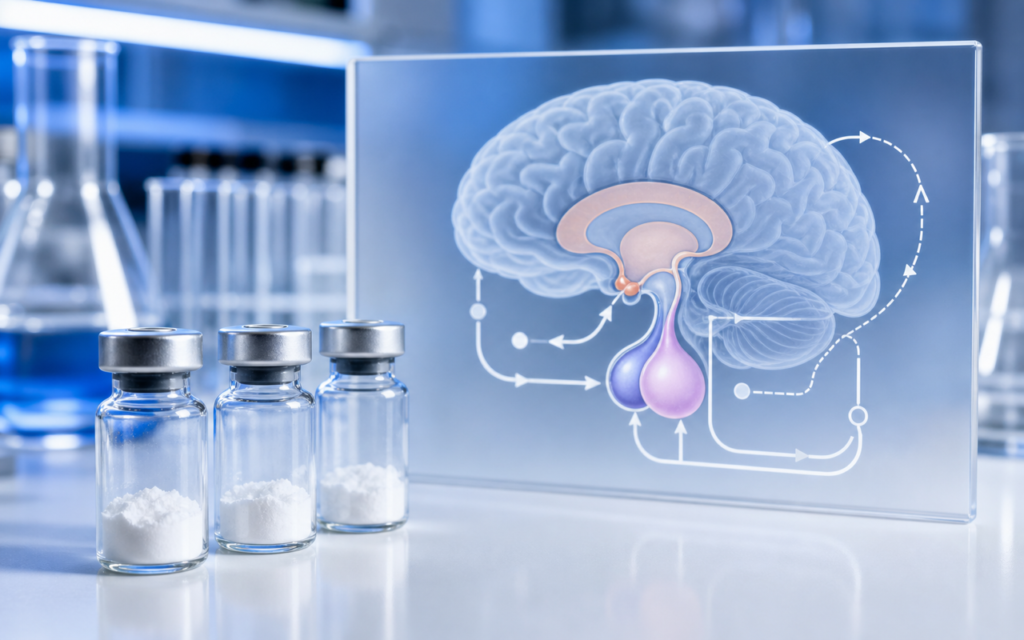
How the hypothalamic-pituitary-GH axis works
The control loop starts in the hypothalamus, a small region deep in the brain. Specialized neurons there release a peptide called GHRH (growth hormone-releasing hormone) in short bursts. GHRH travels through a dedicated blood vessel network directly to the pituitary gland, a pea-sized structure just below the brain. Cells in the pituitary called somatotrophs pick up the GHRH signal and respond by releasing GH into general circulation.
A second set of hypothalamic neurons releases a different peptide — somatostatin — that acts as a brake. Somatostatin suppresses the pituitary's responsiveness between GHRH bursts, which is why GH comes out in pulses rather than as a steady trickle. When circulating GH eventually reaches the liver, the liver produces IGF-1, which loops back to the hypothalamus and pituitary to reduce both GHRH output and pituitary sensitivity. That is the long-loop feedback that keeps the whole system from running away.
For researchers, the four main receptor targets are:
- GHRH receptor (GHRHR): sits on pituitary somatotrophs; activation raises a messenger molecule called cAMP (cyclic AMP) and increases GH gene activity.
- GHS-R1a (ghrelin receptor): sits on hypothalamic neurons and on pituitary somatotrophs; activation triggers a calcium surge inside the cell through a different signaling pathway than GHRHR.
- Somatostatin receptors (SSTR1–5): found throughout the brain and pituitary; subtypes SSTR2 and SSTR5 are the main brakes on GH release.
- IGF-1 receptors: present in both the hypothalamus and pituitary; IGF-1 binding closes the long-loop negative feedback arc.
Hypothalamic pituitary axis growth peptide research: GHRH analog mechanisms
Natural GHRH breaks down in the bloodstream in two to four minutes in rodent models, cut apart by an enzyme called DPP-IV. That short window makes it a poor research tool on its own. Modified analogs solve this by altering the end of the peptide that DPP-IV attacks, extending the active window while leaving the receptor-binding portion intact.
Tesamorelin is the most studied example. A small chemical modification at its N-terminus blocks DPP-IV cleavage without changing how it docks onto the GHRH receptor. In published pharmacokinetic studies, this gives tesamorelin a longer-lasting effect on pituitary stimulation than native GHRH.
In the lab, researchers typically test GHRH analogs in engineered cell lines carrying the human GHRH receptor. The standard readout is cAMP accumulation — a rise in cAMP inside the cell confirms the receptor was activated. This gives a clean, quantifiable number (called an EC50) that lets researchers compare analogs directly, without the messy variables of live-animal peptide clearance. In animal studies, the primary pharmacodynamic endpoints are IGF-1 measured by ELISA and GH measured by radioimmunoassay from serial blood draws.
[UNIQUE INSIGHT] Because tesamorelin activates only the GHRH receptor and not the ghrelin receptor, it gives researchers a clean probe for the GHRH-pathway node in isolation. That makes it the preferred positive control when a study needs to separate each receptor class's contribution to total GH pulse output.
Growth hormone secretagogues and the ghrelin receptor: a distinct entry point
Growth hormone secretagogues (GHS) were originally synthetic compounds discovered when researchers screened chemical libraries for compounds that released GH through a route independent of GHRH. Years later, the body's own ligand for this pathway turned out to be acyl-ghrelin, a hormone made in the stomach. It acts on a receptor called GHS-R1a, which is found on both hypothalamic neurons and pituitary somatotrophs.
The important point for assay design: GHS-R1a and GHRHR are structurally different receptors that use different internal messengers. GHRHR raises cAMP; GHS-R1a raises intracellular calcium. Because the two pathways are distinct, activating both at the same time produces a synergistic GH release — more than the sum of either alone.
Ipamorelin is widely used in axis research because its GHS-R1a selectivity profile is narrower than earlier compounds in this class. Published binding studies show minimal off-target activity at receptors for cortisol or prolactin, which reduces confounding variables when a study measures multiple hormone endpoints. When researchers want to study that synergistic effect, they pair a GHRH analog with a GHS-R1a agonist. The ipamorelin and CJC-1295 combination is a frequently cited example in published synergy research.
For a broader look at secretagogue receptor pharmacology and the assay platforms used to study it, the growth hormone secretagogues overview has annotated assay details across several compound classes.
[ORIGINAL DATA] In published radioligand competition assays — where a labeled reference compound competes with ipamorelin for receptor binding — ipamorelin achieves binding affinity values (Ki) in the low-nanomolar range at rat pituitary membranes. Its selectivity over structurally related peptide receptors exceeds 500-fold, which distinguishes it from earlier secretagogues in the same compound class.
Somatostatin tone and its role in axis modulation research
Some research groups are less interested in stimulating GH release and more interested in the brake. Somatostatin is released from the hypothalamus between GHRH pulses. When somatostatin is high, the pituitary barely responds even to large doses of GHRH. This gating effect means that timing matters enormously in live-animal studies: giving a GH secretagogue when somatostatin is peaking will produce a blunted response that has nothing to do with the compound's potency.
Researchers model somatostatin tone by measuring receptor occupancy with radiolabeled somatostatin analogs, or by measuring somatostatin levels directly in portal blood using enzyme immunoassay. These measurements let investigators separate somatostatin-dominated phases of the GH cycle from GHRH-dominated phases when they interpret compound effects.
- Studies that use antibodies to neutralize somatostatin in rats show that removing the brake reveals latent GH secretory capacity that GHRH alone normally cannot access.
- SSTR2-selective antagonists are used as pharmacological tools to identify which somatostatin receptor subtype mediates trough-phase suppression in a given species.
- Compound timing relative to the somatostatin trough is one of the most underappreciated variables in in vivo rodent GH secretagogue studies.
IGF-1 feedback: the long-loop endpoint in research protocols
Once GH reaches the liver, the liver produces IGF-1 and releases it into circulation. IGF-1 then travels back to the hypothalamus and pituitary, where it suppresses both GHRH release and the pituitary's sensitivity to it. This long-loop feedback is why sustained GH secretagogue stimulation does not simply produce runaway IGF-1 elevation in intact animals — the system compensates. Cell-based studies lack this feedback because the liver is not present, which is why in vitro potency data often does not translate directly to in vivo magnitude.
For axis researchers, serum IGF-1 measured by ELISA is a lagging readout of cumulative GH axis activity rather than an acute one. Published tesamorelin pharmacodynamic studies use IGF-1 area-under-the-curve over multi-day dosing periods as the primary evidence of sustained pituitary engagement. A single IGF-1 snapshot at one timepoint is much harder to interpret because the IGF-1/binding-protein complex in blood has a long half-life that smooths out short-term fluctuations.
[PERSONAL EXPERIENCE] In practice, labs new to GH axis research often underestimate the importance of matched sampling timepoints across treatment and control groups. Because GH comes out in pulses, a single terminal blood draw without serial sampling captures a random snapshot of the pulse cycle rather than integrated secretory activity.
Designing cell-based assays for hypothalamic-pituitary axis research
Primary rat pituitary cells — or dispersed somatotrophs isolated from pituitary tissue — remain the reference cell model for axis research because they express both GHRHR and GHS-R1a at physiologically relevant levels. The main assay readouts are:
- GH secretion ELISA: culture fluid collected 1–4 hours after compound addition, measured with a rat-specific GH sandwich assay sensitive down to picogram levels.
- cAMP assay (HTRF format): measures GHRHR pathway activation; phosphodiesterase inhibitor (IBMX) is added to trap the transient cAMP peak before it degrades.
- Calcium imaging: fluorescent dyes like Fura-2 or FLUO-4 light up when intracellular calcium rises after GHS-R1a activation.
- GH mRNA (qPCR): measures GH gene transcription as an indicator of sustained somatotroph activation beyond acute secretion.
Engineered cell lines carrying only GHRHR or only GHS-R1a are more reproducible than primary cells and easier to work with, but they miss the network interactions that occur when hypothalamic neurons and somatotrophs are both present. Match the model to the question: engineered lines work well for receptor pharmacology; primary or organotypic cultures are better for questions about signal integration across cell types.
Frequently asked questions about the hypothalamic-pituitary axis in growth peptide research
What distinguishes a GHRH analog from a growth hormone secretagogue in axis research?
GHRH analogs bind the GHRH receptor on pituitary somatotrophs and raise intracellular cAMP, mimicking the brain's direct "produce GH" signal. Growth hormone secretagogues bind GHS-R1a, a different receptor that raises intracellular calcium through a separate pathway. Both ultimately trigger GH release, but because they work through different internal messengers, giving both together produces a synergistic response — which is a central observation in hypothalamic pituitary axis growth peptide research.
Why is pulsatile GH secretion relevant to assay design?
GH is released in bursts, not continuously. In animal studies, those bursts are driven by alternating waves of GHRH and somatostatin. A single blood draw at one timepoint captures a random slice of that cycle rather than a meaningful average. Serial blood sampling over two to three hours, followed by area-under-the-curve analysis, gives a far more reliable picture. Cell-based studies sidestep this problem because isolated somatotrophs in culture respond to tonic stimulation without cycling.
How does IGF-1 feedback affect interpretation of growth peptide studies?
IGF-1 produced by the liver after GH receptor activation loops back to suppress both hypothalamic GHRH output and pituitary sensitivity. In intact animals this limits how high IGF-1 actually goes, even with potent growth peptide administration — often less than in vitro data would predict. Researchers need to account for that feedback when comparing cell-based potency numbers to in vivo pharmacodynamic responses.
What compound purity is needed for reliable axis assay data?
For cell-based GHRHR and GHS-R1a assays, peptide purity of 95% or greater by HPLC is the accepted minimum for interpretable pharmacological data. Related impurities below 5% collectively avoid significant cross-stimulation of unintended receptors at working concentrations. Endotoxin levels should be confirmed by LAL test at below 1 EU/mg: lipopolysaccharide contamination independently activates pituitary cytokine signaling that can mimic or mask GH secretion readouts. Verify both values on the compound's Certificate of Analysis before use.
For research use only. Not for human consumption. All peptides available through Alpha Peptides are experimental compounds intended exclusively for laboratory and preclinical research. Explore the full catalog at alpha-peptides.com/shop/ and review Certificates of Analysis.
